Gene Therapy for Post-Treatment Lyme Disease Syndrome
Madelyn Conn, Savannah Avery, Amrit Bhatia, Ahmed Ali Alnamos
Last updated: August 25, 2021
-
A. Abstract
Post-treatment Lyme Disease Syndrome (PTLDS) is a multi-organ animal-borne disease caused by an antibiotic-resistant strain of the bacteria Borrelia burgdorferi (Bb) referred to as Bb hereafter. The disease is transmitted through tick bites; however, it is originally contracted from wild mammals, especially deer and small rodents. After infection the bacteria travels into the bloodstream and attaches onto fibronectin of the extracellular matrix, further spreading the bacteria throughout the entire body. PTLDS affects 10% of people infected with Lyme Disease and is described by more serious symptoms as time progresses.31 Some of the more dire symptoms of PTLDS include arthritis, facial palsy, heart palpitations, inflammation of the brain and spinal cord, and nerve pain.21 To combat these symptoms, we aim to identify some promising antigens that are expressed on Bb outer surface, then to engineer Chimeric Antigen Receptor (CAR) T-cells to eliminate the bacteria. Finally, CAR T-cells can be injected into a patient after their efficiency has been verified in vitro and in vivo using the most accurate animal models available in the market. This proposal is an attempt to establish an official treatment protocol for PTLDS to relieve its symptoms and offer solace to patients who have lived with this disease for years with no current cure or standard treatment method.21
-
B. Specific aims
1. Identify Outer surface proteins (Osps) and antigens expressed on Bb
2. Build CAR T-cells that bind to the identified receptors to eliminate Bb
3. Test the efficiency of the Built CAR T-cells in vitro and in vivo using indicative models
-
C. Introduction
Lyme Disease is a multi-organ animal-borne disease caused by the bacteria Bb. The infection is transmitted via ticks, and a human is infected through a tick bite to the skin.1 The tick must feed for approximately 36 hours before the bacteria fully transmits itself.2 There are several clinical features of this disease, including fever, chills, myalgias, fatigue, headache, and malaise, which may precede onset of a characteristic “Erythema migrans” rash by a few days.21 These symptoms generally appear 7 to 14 days after the bacterial transmission; however, it is possible that these symptoms may be offset and appear up to a month after contact with the tick.2 Generally, treatment of the bacterial infection is done by prescribing oral doses of doxycycline, amoxicillin, or other common antibiotics. This directive should be followed and completed over the course of 14 to 21 days.2 However, cases have been seen where simple antibiotic treatment plans only temporarily minimize symptoms.17 Furthermore, Bb has developed various immune evasion mechanisms that may render antibody responses ineffective, therefore supporting ongoing infection. There are numerous documentations of up-regulation or down-regulation responses of immunogenic surface proteins during infection that make it harder to battle the infection.15 This allows the disease to manifest into Post-treatment Lyme Disease Syndrome, more commonly known, although clinically rejected, as “Chronic Lyme Disease.”31 Because antibiotic drug delivery can be difficult, the bacteria can manifest throughout the body by using the surface lipoprotein, BBK32, to bind to fibronectin, forming endothelial interactions as it travels along the bloodstream.25 Previous studies have been done to engineer Bb so that it expresses green fluorescent proteins (GFPs) and these interactions and adhesions were able to be tracked.24 However, further damage is done when the protein binds to glycosaminoglycans (GAGs), as they are also ubiquitous on cell surfaces.8, 25 GAG binding is independent from fibronectin binding and causes different effects.8 However, it was determined that even when BBK32 was knocked-out, the bacteria was still able to spread.18 This suggests that the bacteria has other proteins to support its movement within the body. The body has many natural defense mechanisms. The primary line of defense in the bloodstream is composed of leukocytes, or white blood cells. There are five major types of white blood cells, including neutrophils, lymphocytes, monocytes, eosinophils, and basophils. Lymphocytes are produced by bone marrow and differentiate into B-cells (stay in bone marrow) and T-cells (travel to the thymus for specialization). 22 B-cells and T-cells can fall into two main categories: effector cells that are activated by antigens to fight an active infection, or memory cells that recognize past infections and “remember” how to respond to and overcome them. 22 Specifically, T-cells are further divided into three types of specific cells: cytotoxic, helper, and regulatory T-cells. 22 Cytotoxic T-cells, often referred to as killer T-cells, destroy infected or foreign cells in the body.22 Helper T-cells communicate with other T-cells and B-cells and direct their immune response.22 Regulatory T-cells suppress the immune system when necessary, keeping immune response in check.22
-
D. Proposed research
Our first step is to identify the most important antigens that are expressed on the outer surface of the Bb. Bb has some principal genes that enhance its resistance against reactive oxygen and nitrogen species such as BB0164 and BB0219.27 The latter encodes for Borrelia manganese transporter A (BmtA) which is required for virulence of Bb spirochete to infect and survive in its arthropod vector and is essential for mammalian infectivity.35 Bb can be significantly compromised under conditions of increased oxidative stress due to a reduced energy flux and contribute to a defect in the colonization of the mouse model of Lyme disease. The levels of Decorin binding protein A (DbpA), BBK32, and Outer surface protein C (OspC) are reduced in the sodA mutant compared to the control strains.7 The DbpBA deletion strain showed no detectable spirochaetes except immediately following inoculation for mice infected with 105 organisms. At no other time point are spirochaetes visible in Bb deleted for DbpBA. The borrelial numbers for the BBK32 mutant are approximately 20-fold lower at days 4 and 7 in the lymph node tissue relative to the infectious parent and remained significantly lower even at later time points.12 BB0347 is a fibronectin binding protein expressed on the surface of Bb, which appears to be specific to the joint like BBK32. Bb expresses surface proteins that recruit host complement regulatory factors to the bacterial surface. Bb achieves resistance to the alternative component cascade active in the blood of the host. This increased blood burden is reduced upon the elimination of the GAG binding domain of BBK32. Bb burdens in the bloodstream are increased upon restoration of OspC expression.4 OspC type K (RST2) strains are most often found in the joints of patients with Lyme arthritis, but RST1 strains are most frequent in those with antibiotic refractory arthritis.14 While Bb requires addition of GlcNAc for cell wall synthesis, the carbohydrate can also serve as an energy source; Bb expresses transporters and enzymes for utilization of some carbohydrates.33 Antibodies directed against OspA readily killed the spirochetes.16 During late disseminated infection, Bb colonizes multiple regions of the dura mater, and are predominantly found adhering to blood vessels. The increase in the number of T-cells in the meninges is suggestive of adaptive immune response to Bb colonization of the dura mater.5 Ingested Bb is a potent inducer of apoptosis in monocytes, and Bb isolates are resistant to various components of the human innate immune response.23 OspA and OspC are well-characterized ligands for Toll-Like Receptor 2 (TLR2). TLR2 and TLR7 signaling contribute to the production of NF-kappaB-dependent cytokines by human peripheral blood mononuclear cells (PBMCs) in response to live Bb.20 Mutations of the hrpA gene obliterate Bb infectivity.29 The hrpA-D126A and hrpA-E127A mutations (Motif II, ATP binding and hydrolysis) resulted in a complete loss of Bb infectivity in wild-type mice like the hrpA knockout strain.29 Bb p83/100 antigen protein is known to elicit a strong IgM and IgG response in all stages of Lyme disease. The Borrelia persistence protein A (BptA) is conserved in all the sensu lato strains and is an Osp as well. All diderms can be characterized by the presence of beta-barrel assembly machinery (BAM complex) which is in the outer membrane.32
The second step is to leverage the new therapeutic method that has been effective for blood cancer since both diseases manifest in the bloodstream. The use of Chimeric Antigen Receptor (CAR) T-cells is based on the fundamental concept that T-cells use receptors to identify antigens on cells in the body and use this to destroy cells that do not belong. CAR T-cells are T-cells taken from an individual which are genetically modified to identify an antigen that original T-cells did not recognize as a threat. Once modified and tested they are inserted back into a patient where they are able to eradicate diseases that previously had no cure. Our goal is to identify the sequences of the antigens expressed on the outer surface of Bb and obtain T-cells from the patient. Then, we will modify the genetic code of the patient’s T-cells in vitro using a lentivirus to recognize the Bb antigens. Preferably, we will design unique CAR T-cells for each important Bb antigen to test for the most effective one. After testing the CAR T-cells efficiency in vitro and in vivo using predictive animal models. CAR T-cells can be injected into the patient where they will recognize Bb, duplicate, then destroy and survive in the patient’s body.
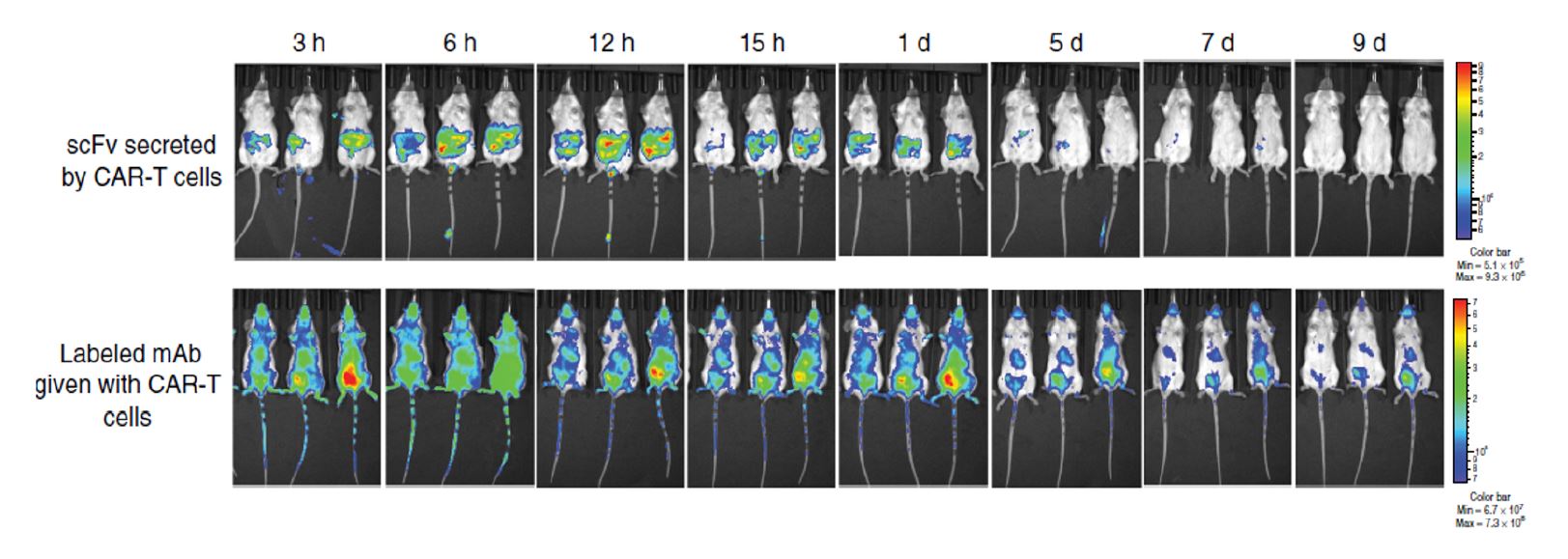
Figure 2. Imaging and Quantification CAR T-Cells Efficiency30
-
E. Anticipated problems / alternative methods
A problem that could arise may have to do with the specific antigens that are targeted. If the antigens which are the target of the CAR T-cells are also present on the cells of certain tissues in the body, then the CAR T-cells may target the tissue cells unintentionally which may lead to damage and other complications. One way to prevent this would be to cross reference the proposed antigens against a theoretical library of all the antigens in the human body. This theoretical library could be used to find which antigens are both effective and safe to target on the bacteria. There are some alternative methods to the proposal discussed in this document. One is to use CRISPR-Cas9 to target the gene that codes for RNA-polymerase in the bacteria, thus hindering functionality. A second approach would be to use an integral carbohydrate, GlcNAc, which is utilized for the synthesis of the bacteria’s cell membrane as well as for energy production. If a link was attached to GlcNAc that an siRNA or an sgRNA could be attached to, then the siRNA could enter the bacteria and silence critical genes for the bacteria’s survival and functionality, or sgRNA could guide the Bb onboard Cas9 to delete the Bb RNA polymerase coding gene. A third approach could be to design a ligand that binds to the manganese transporter of the bacterial cell. Currently, when the cell is under attack from outside agents, it recruits manganese which enters the cell and negates the attacks. If the transporter for the manganese was blocked by a ligand then that could open the bacterial cell to other attacks. A final alternative would be to completely stop the spread of PTLDS by killing off every tick in the world. Ticks are the only known infecting agents of the bacteria and thus if there were no ticks to be carriers then the bacteria would not spread and there would be no new cases of Lyme Disease. However, this method has innumerable repercussions as one cannot predict the impact that removing an entire stratum from the food chain could have.
-
References
1. Biesiada, G., Czepiel, J., Leśniak, M. R., Garlicki, A., & Mach, T. (2012). Lyme disease: review. Archives of medical science : AMS, 8(6), 978–982. doi:10.5114/aoms.2012.30948
2. Bratton, R. L., Whiteside, J. W., Hovan, M. J., Engle, R. L., & Edwards, F. D. (2008). Diagnosis and Treatment of Lyme Disease. Mayo Clinic Proceedings, 83(5), 566-571. doi:https://doi.org/10.4065/83.5.566
3. Brown, & Dennis. (2017). Borrelia burgdorferi infection induces lipid mediator production during Lyme arthritis. Biochimie, 141, 86-90.
4. Caine, J., & Coburn, J. (2015). A short-term Borrelia burgdorferi infection model identifies tissue tropisms and bloodstream survival conferred by adhesion proteins. Infection and Immunity, 83(8), 3184-3194.
5. Divan, Ali, Casselli, Timothy, Narayanan, S. Anand, Mukherjee, Sanjib, Zawieja, David C., Watt, John A., . . . Newell-Rogers, M. Karen. (2018). Borrelia burgdorferi adhere to blood vessels in the dura mater and are associated with increased meningeal T-cells during murine disseminated borreliosis. PLoS ONE,13(5), E0196893.
6. Ebady, Niddam, Boczula, Kim, Gupta, Tang, . . . Moriarty. (2016). Biomechanics of Borrelia burgdorferi Vascular Interactions. Cell Reports, 16(10), 2593-2604.
7. Esteve-Gassent, M., Smith, T., Small, C., Thomas, D., & Seshu, J. (2015). Absence of sodA Increases the Levels of Oxidation of Key Metabolic Determinants of Borrelia burgdorferi: E0136707. PLoS ONE, 10(8), E0136707.
8. Fischer, J. R., LeBlanc, K. T., & Leong, J. M. (2006). Fibronectin binding protein BBK32 of the Lyme disease spirochete promotes bacterial attachment to glycosaminoglycans. Infection and immunity, 74(1), 435–441. doi:10.1128/IAI.74.1.435-441.2006
9. Garcia, B., Zhi, H., Hook, B., & Skare, J. (2016). Borrelia burgdorferi BBK32 inhibits the classical pathway by blocking activation of the C1 complement complex. PLoS Pathogens, 12(1), E1005404.
10. Grube, Blanche D., & Douglas, Leslie J. (2017). How finding a Lyme spirochete in a root-canal tooth led to the development of a new test panel. Townsend Letter, (408), 95.
11. Harris, G., Ma, W., Maurer, L., Potts, J., & Mosher, D. (2014). Borrelia burgdorferi protein BBK32 binds to soluble fibronectin via the N-terminal 70-kDa region, causing fibronectin to undergo conformational extension. The Journal of Biological Chemistry, 289(32), 22490-22499.
12. Hyde, J., Weening, E., Chang, M., Trzeciakowski, J., Höök, M., Cirillo, J., & Skare, J. (2011). Bioluminescent imaging of Borrelia burgdorferi in vivo demonstrates that the fibronectin‐binding protein BBK32 is required for optimal infectivity. Molecular Microbiology, 82(1), 99-113.
13. Jabbari, Neda, Glusman, Gustavo, Joesch-Cohen, Lena M., Reddy, Panga Jaipal, Moritz, Robert L., Hood, Leroy, & Lausted, Christopher G. (2018). Whole genome sequence and comparative analysis of Borrelia burgdorferi MM1. PLoS ONE, 13(6), E0198135.
14. Jones, K., McHugh, G., Glickstein, L., & Steere, A. (2009). Analysis of Borrelia burgdorferi genotypes in patients with lyme arthritis: High frequency of ribosomal RNA intergenic spacer type 1 strains in antibiotic‐refractory arthritis. Arthritis & Rheumatism, 60(7), 2174-2182.
15. Kenedy, M. R., Lenhart, T. R., & Akins, D. R. (2012). The role of Borrelia burgdorferi outer surface proteins. FEMS immunology and medical microbiology, 66(1), 1–19. doi:10.1111/j.1574-695X.2012.00980.x
16. Kung, F., Kaur, S., Smith, A., Yang, X., Wilder, C., Sharma, K., . . . Pal, U. (2016). A Borrelia burgdorferi Surface-Exposed Transmembrane Protein Lacking Detectable Immune Responses Supports Pathogen Persistence and Constitutes a Vaccine Target. The Journal of Infectious Diseases, 213(11), 1786-1795.
17. Lacout, A., El Hajjam, M., Marcy, P. Y., & Perronne, C. (2018). The Persistent Lyme Disease: "True Chronic Lyme Disease" rather than "Post-treatment Lyme Disease Syndrome". Journal of global infectious diseases, 10(3), 170–171. doi:10.4103/jgid.jgid_152_17
18. Li, X., Liu, X., Beck, D. S., Kantor, F. S., & Fikrig, E. (2006). Borrelia burgdorferi Lacking BBK32, a Fibronectin-Binding Protein, Retains Full Pathogenicity. Infection and Immunity, 74(6), 3305. doi:10.1128/IAI.02035-05
19. Lin, Y., Chen, Q., Ritchie, J., Dufour, N., Fischer, J., Coburn, J., & Leong, J. (2015). Glycosaminoglycan binding by Borrelia burgdorferi adhesin BBK32 specifically and uniquely promotes joint colonization. Cellular Microbiology, 17(6), 860-875.
20. Love, A., Schwartz, I., & Petzke, M. (2014). Borrelia burgdorferi RNA Induces Type I and III Interferons via Toll-Like Receptor 7 and Contributes to Production of NF-kB-Dependent Cytokines. Infection and Immunity, 82(6), 2405-2416.
21. Lyme Disease | Lyme Disease | CDC. Centers for Disease Control and Prevention. https://www.cdc.gov/lyme/index.html. Published December 21, 2018. Accessed April 17, 2019.
22. Lymphocytes: Definition, Counts, and More. Healthline. https://www.healthline.com/health/lymphocytes#function. Accessed April 17, 2019.
23. Mason, Herkes, Krupna-Gaylord, Oei, Poll, Wormser, . . . Hovius. (2015). Borrelia burgdorferi clinical isolates induce human innate immune responses that are not dependent on genotype. Immunobiology,220(10), 1141-1150.
24. Moriarty, T. J., Norman, M. U., Colarusso, P., Bankhead, T., Kubes, P., & Chaconas, G. (2008). Real-time high resolution 3D imaging of the lyme disease spirochete adhering to and escaping from the vasculature of a living host. PLoS pathogens, 4(6), e1000090. doi:10.1371/journal.ppat.1000090
25. Niddam, A. F., Ebady, R., Bansal, A., Koehler, A., Hinz, B., & Moriarty, T. J. (2017). Plasma fibronectin stabilizes Borrelia burgdorferi endothelial interactions under vascular shear stress by a catch-bond mechanism. Proceedings of the National Academy of Sciences, 114(17), E3490. doi:10.1073/pnas.1615007114
26. Petzke, M., Iyer, R., Love, A., Spieler, Z., Brooks, A., & Schwartz, I. (2016). Borrelia burgdorferi induces a type I interferon response during early stages of disseminated infection in mice. BMC Microbiology,16(29), 29.
27. Ramsey, Meghan E., Hyde, Jenny A., Medina-Perez, Diana N., Lin, Tao, Gao, Lihui, Lundt, Maureen E., . . . Hu, Linden T. (2017). A high-throughput genetic screen identifies previously uncharacterized Borrelia burgdorferi genes important for resistance against reactive oxygen and nitrogen species. PLoS Pathogens, 13(2), E1006225.
28. Salman-Dilgimen, A., Hardy, P., Dresser, A., Chaconas, G., & Adler, B. (2011). HrpA, a DEAH-Box RNA Helicase, Is Involved in Global Gene Regulation in the Lyme Disease Spirochete (HrpA Is Involved in Global Gene Regulation). PLoS ONE, 6(7), E22168.
29. Salman-Dilgimen, A., Hardy, P., Radolf, J., Caimano, M., & Chaconas, G. (2013). HrpA, an RNA Helicase Involved in RNA Processing, Is Required for Mouse Infectivity and Tick Transmission of the Lyme Disease Spirochete. PLoS Pathogens, 9(12), E1003841.
30. Sarwish Rafiq, Oladapo O Yeku, Hollie J Jackson, Terence J Purdon, Dayenne G Van Leeuwen, Dylan J Drakes, . . . Renier J Brentjens. (2018). Targeted delivery of a PD-1-blocking scFv by CAR T-cells enhances anti-tumor efficacy in vivo. Nature Biotechnology, 36(9), 847-856.
31. Ścieszka J, Dąbek J, Cieślik P. Post-Lyme disease syndrome. Reumatologia. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4847307/. Published April 10, 2015. Accessed April 17, 2019.
32. Verhaegh D, Joosten LAB, Oosting M. The role of host immune cells and Borrelia burgdorferi antigens in the etiology of Lyme disease. Eur. Cytokine Netw. 2017; 28(2) : 70-84 doi:10.1684/ecn.2017.0396
33. Von Lackum, K., & Stevenson, B. (2005). Carbohydrate utilization by the Lyme borreliosis spirochete, Borrelia burgdorferi. FEMS Microbiology Letters, 243(1), 173-179.
34. Zhao, Chen, Francisco, Zhang, & Wu. (2018). The application of CAR T-cell therapy in hematological malignancies: Advantages and challenges. Acta Pharmaceutica Sinica B, 8(4), 539-551.
35. Zhiming Ouyang, Ming He, Tara Oman, X. Frank Yang, & Michael V. Norgard. (2009). A manganese transporter, BB0219 (BmtA), is required for virulence by the Lyme disease spirochete, Borrelia burgdorferi. Proceedings of the National Academy of Sciences, 106(9), 3449-3454.